

Różnica między cykliczną i niecykliczną fotofosforylacją
- 1421
- 222
- Emilia Rudziński
Większość materiałów organicznych wymaganych przez organizmy powstaje z produktów fotosyntezy. Fotosynteza obejmuje konwersję energii światła w energię, która może być wykorzystywana przez komórkę, w szczególności energię chemiczną. W roślinach i glonach fotosynteza występuje w organelle zwanej chloroplastem, która zawiera błonę zewnętrzną, błonę wewnętrzną i błonę tylakoidową (https: // en.Wikipedia.org/wiki/chloroplast).
Fotosyntezy można podzielić na dwie główne części: (1) fotosyntetyczne reakcje transferu elektronów („reakcje światła”) i (2) reakcje utrwalania węgla („reakcje ciemne”). „Reakcje lekkie” obejmują energetyzujące światło słoneczne elektrony w fotosyntetycznym chlorofilu pigmentu, który następnie przemieszczają się wzdłuż łańcucha transportu elektronów w błonie tylakoidowej, co powoduje powstawanie ATP i NADPH. „Mroczne reakcje” obejmują wytwarzanie związków organicznych z CO2 przy użyciu ATP i NADPH wytwarzanych przez „reakcje świetlne” i nie zostaną omówione dalej w tym artykule.
Fotosynteza obejmuje użycie dwóch fotosystemów (Photosystem I i Photosystem II) Aby wykorzystać energię światła za pomocą elektronów do wytwarzania ATP i NADPH, które później mogą być wykorzystywane przez komórkę jako energia chemiczna do wytwarzania związków organicznych. Scenariusz to duże kompleksy białkowe, które specjalizują się w gromadzeniu energii świetlnej i przekształcaniu jej w energię chemiczną. Strakiery fotograficzne składają się z dwóch części: kompleksu antenowego i centrum reakcji fotochemicznej. Kompleks anteny jest ważny w przechwytywaniu energii światła i przekazywaniu tej energii do fotochemicznego centrum reakcji, co następnie przekształca energię w użyteczne formy dla komórki.
Po pierwsze, światło podnieca elektron w cząsteczce chlorofilu w kompleksie anteny. Obejmuje to foton światła powodującego przemieszczenie elektronu na orbital o wyższej energii. Gdy elektron w cząsteczce chlorofilu jest wzbudzony, jest on niestabilny na orbicie wyższej energii, a energia jest szybko przenoszona z jednej cząsteczki chlorofilu do drugiej przez przenoszenie energii rezonansowej, aż osiągnie cząsteczki chlorofilu w obszarze znanym jako Centrum reakcji fotochemicznej. Stąd wzbudzone elektrony są przekazywane do łańcucha akceptorów elektronów. Energia światła powoduje przeniesienie elektronów ze słabego dawcy elektronów (o silnym powinowactwie do elektronów) do silnego dawcy elektronów w jego zmniejszonej postaci (przewożącego elektron wysokoenergetyczny). Konkretne dawcy elektronów używane przez dany organizm lub system fotosystemowy mogą się różnić i zostaną omówione poniżej dla fotosystemów I i II w roślinach.
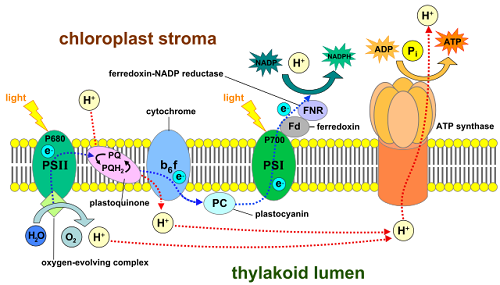
W roślinach fotosynteza powoduje produkcję ATP i NADPH przez dwuetapowy proces znany jako Niecykliczna fotofosforylacja. Pierwszy etap niecyklicznej fotofosforylacji obejmuje fotosystem II. Elektrony wysokoenergetyczne (spowodowane energią świetlną) z cząsteczek chlorofilu w centrum reakcji Photosystem II są przenoszone na cząsteczki chinonu (silne dawcy elektronów). Photosystem II wykorzystuje wodę jako słabego dawcy elektronów w celu zastąpienia niedoborów elektronów spowodowanych przeniesieniem elektronów wysokoenergetycznych z cząsteczek chlorofilu do cząsteczek chinonu. Odbywa się to przez enzym rozdzierający wodę, który umożliwia usuwanie elektronów z cząsteczek wody w celu wymiany elektronów przeniesionych z cząsteczki chlorofilu. Gdy 4 elektrony są usuwane z dwóch cząsteczek H2O (odpowiadających 4 fotonom), O2 jest uwalniany. Zmniejszone cząsteczki chinonu następnie przekazują elektrony wysokoenergetyczne do pompy protonowej (H+) znanej jako cytochrom B6-F złożony. Cytochrom B6-F złożone pompy H+ w przestrzeni tylakoidów, tworząc gradient stężenia przez błonę tylakoidową.
Ten gradient protonu napędza następnie syntezę ATP przez syntazę ATP enzymu (zwaną także ATPazą F0F1). Syntaza ATP stanowi środki dla jonów H+ do podróży przez błonę tylakoidową, w dół ich gradientu stężenia. Ruch jonów H+ w dół ich gradientu stężenia napędza tworzenie ATP z ADP i PI (fosforanu nieorganicznego) przez syntazę ATP. Syntaza ATP występuje w bakteriach, archea, roślinach, algach i komórkach zwierzęcych i odgrywa rolę zarówno w oddychaniu, jak i fotosyntezie (https: // en.Wikipedia.org/wiki/atp_syntaza).
Ostatnim transferem elektronów fotosystemu II jest przeniesienie elektronów do cząsteczki chlorofilu z niedoborem elektronów w centrum reakcji fotosystemu i. Podekscytowany elektron (spowodowany energią świetlną) z cząsteczki chlorofilu w centrum reakcji fotosystemu i jest przenoszony na cząsteczkę zwaną ferredoksyną. Stamtąd elektron jest przenoszony do NADP+, aby utworzyć NADPH.
Niecykliczna fotofosforylacja wytwarza 1 cząsteczkę ATP i 1 cząsteczkę NADPH na parę elektronów; Jednak utrwalenie węgla wymaga 1.5 cząsteczek ATP na cząsteczkę NADPH. Aby rozwiązać ten problem i wyprodukować więcej cząsteczek ATP, niektóre gatunki roślin używają procesu znanego jako Cykliczna fotofosforylacja. Cykliczna fotofosforylacja obejmuje tylko fotosystem I, a nie fotosystem II i nie tworzy NADPH ani O2. W cyklicznej fosforylacji elektrony wysokoenergetyczne z fotosystemu I są przenoszone do cytochromu B6-F złożone zamiast przenieść do NADP+. Elektrony tracą energię, gdy przechodzą przez cytochrom B6-F W rezultacie złożone z powrotem do chlorofilu fotosystemu I i H+ jest pompowane przez błonę tylakoidową. Zwiększa to stężenie H+ w przestrzeni tylakoidowej, która napędza produkcję ATP przez syntazę ATP.
Poziom fotofosforylacji niecyklicznej i cyklicznej, która występuje w danej komórce fotosyntetycznej, jest regulowany na podstawie potrzeb komórki. W ten sposób komórka może kontrolować, ile energii światła przekształca w moc zmniejszającą (napędzaną przez NADPH) i ile jest przekształcane w wiązania fosforanowe o wysokiej energii (ATP).