

Różnica między protistami a grzybami

- 3040
- 955
- Prokop Cebula
Wstęp
Protist i grzyby obejmują dwa wyjątkowe królestwa życia. Protist wykazują solidną wariancję cech komplikujących ich taksonomię. Grzyby są znacznie prostsze do scharakteryzowania. Grzyby odbiegały od protistów około 1.5 miliardów lat temu [1] zdarzenie, które pociągało za sobą utratę flagów podczas przejścia od siedliska wodnego do naziemnego z towarzyszącym rozwojem nowych mechanizmów rozprzestrzeniania się zarodników opartych na opartych. Grzyby dostosowały rozpoznawalny zestaw cech, które pomagają wyjaśnić ich różnice od protistów.
Różnice komórkowe
Protist to organizmy jednokomórkowe. Większość grzybów jest wielokomórkowa i ustrukturyzowana w rozgałęzionym wydłużonym układowi nitkowatnim strzępek [1]. Odgałęzione struktury strzępek składają się z jednej lub (zwykle) większej liczby komórek zamkniętych w rurowej ścianie komórkowej [1]. Większość protistów ma kształt sferyczny, który jest nieoptymalny dla uzyskiwania tlenu przez dyfuzję. Duże protiści mają wydłużony kształt, aby zaspokoić zwiększoną potrzebę dyfuzji tlenu [3].
Rozmiar komórki
Protisty jednokomórkowe są w większości mikroskopijne, ale rzadkie przykłady znaleziono tysiące metrów kwadratowych w obszarze [3]. Grzyby są zwykle wystarczająco duże, aby można je zaobserwować gołym okiem, ale istnieje duża liczba gatunków mikroskopowych [1].
Błona komórkowa
Protist mogą zawierać podobne do roślin ściany komórkowe, zwierzęce ściany komórkowe, a nawet peleryny zapewniające ochronę przed środowiskiem zewnętrznym [3]. Wielu protistów nie ma ściany komórkowej [3]. W przeciwieństwie do odmiany błony komórkowej protisty, decydującą cechą grzybów jest wszechobecna obecność chitynowej ściany komórkowej [14].
Organizacja wewnątrzkomórkowa
Grzyby składają się z skomplikowanego systemu strzępek podzielonych przez system podziału SEPTA [1]. SEPTA nie znaleziono u żadnych protistów [3]. Grzybicze przegrody dzielą strzępki na przepuszczalne przedziały [1]. Perforacja przegrody umożliwia translokację organelli, w tym rybosomów, mitochondriów i jąder między komórkami [3]. Protist organelle istnieją w nieistotnej cytoplazmie [3].
Przypadki komórkowe
W przeciwieństwie do głównie stacjonarnych grzybów, protistów są ruchliwe [1,3], a ruchliwość ta odróżnia protistów morfologicznie od grzybów poprzez dodanie dodatków komórkowych. Protisty często zawierają dodatki, takie jak rzęsy, flagella i pseudopodia [3]. Grzyby na ogół nie mają przydatków komórkowych, choć istnieją rzadkie przykłady dbałówek konidialnych w grzybach [4].
Oddychanie
Protist oddychanie
i) Protist Aerobic Respiration
Protisty uzyskują tlen przez dyfuzję, co ogranicza ich zdolność do wzrostu komórkowego [3]. Niektórzy protisti, takie jak fitoflagelaty, przeprowadzają zarówno autotroficzny, jak i oksydacyjny heterotroficzny metabolizm [3]. Protist metabolizm działa optymalnie poprzez szeroki zakres temperatur i ilości zużycia tlenu. Jest to produkt uboczny mnóstwa nisz, które zamieszkują, które mają ogromny zakres temperatur i dostępności tlenu [3].
ii) Protist beztlenowy oddychanie
Obowiązkowe oddychanie beztlenowe istnieje wśród pasożytniczych protistów, rzadkość dla eukariotów [3]. Wielu obowiązkowych protistów anaerobowych brakuje oksydazy cytochromu, co powoduje atypowe mitochondria [3].
iii) oddychanie grzybowe
Większość grzybów oddychania aerobowo przy użyciu rozgałęzionych łańcuchów oddechowych do przenoszenia elektronów z NADH do tlenu [5]. Dehydrogenazy NADH grzybowej stosuje się do katalizowania utleniania matrycy NADH i są zdolne do tego, nawet w obecności niektórych inhibitorów, takich jak rotenon [5]. Grzyby wykorzystują również alternatywne oksydazy do oddychania w obecności inhibitorów ubichinolu: oksydoreduktazy cytochromu C i oksydazy cytochromu C [5]. Alternatywne oksydazy prawdopodobnie umożliwiają skuteczną patogenność w obecności mechanizmów obrony opartej na tlenku tlenku [5].
Osmoregulacja
Protist, który zamieszkuje środowisko wodne, mają wzmocnienie struktur komórkowych nie znalezionych w grzybach. Ta wzmocnienie umożliwia wyższy stopień osmoregulacji. Wakuole skurczowe to protistyczne organelle, które umożliwiają osmoregulację i zapobiegają pęcznieniu i pęknięciu komórek [3]. Wakuole skurczowe są otoczone systemem kanalików i pęcherzyków zbiorowo zwanych gąbką, który pomaga w wydaleniu wakuoli skurczowej z komórki [3]. Wakuole skurczowe są znacznie mniej obfite w grzybach [1,3].
Różnice mitochondrialne
Protist mitochondrialne genomy
W przeciwieństwie do grzybów, genomy mitochondrialne (MT) zachowały szereg przodków proto-mitochondryjskich elementów genomowych. Jest to widoczne przez redukcję genów w grzybach [6]. Protist Mtgenomes waha się od genomu 6kb Plasmodium falciparum do genomu 77 kb choanoflagellanu Monosiga brevicollis, mniejszy zasięg niż grzyby [6]. Średnia protistyczna wielkość MTGenome ma 40 kb znacznie mniejszą niż średnia wielkość genomu mitochondrialnego grzybowego [6].
Protist Mtgenomes są kompaktowe, bogate w ekson i często składają się z nakładających się regionów kodujących [6]. Niekodująca przestrzeń intronowa stanowi mniej niż 10% całkowitego protystowego rozmiaru MTGenome [6]. Duża część protist mtDNA nie ma intronów grupy I ani grupy II [6]. Zawartość A+T jest wyższa w protist MTGenomes w porównaniu do grzybów [6].Zawartość genów protist mtgenomes przypomina mtgenomy roślin bardziej niż grzybowe mtgenomy [6]. W przeciwieństwie do grzybów, protist MTGenomes koduje zarówno dla dużych, jak i małych podjednostek RNA [6].
Genomy mitochondrialne grzybowe
Grzyby ewoluowały z protistów, a ich rozbieżność charakteryzuje się redukcją genów i dodaniem intronu [6]. W porównaniu z bogatymi genami protistycznymi mtgenomami, grzybowe mtgenomy zawierają mnóstwo regionów międzygenowych składających się z niekodujących powtórzeń i intronów, które są głównie intronami grupy I [7]. Zróżnicowanie wielkości grzybów MTGenome jest w większości wyjaśniane przez regiony intronowe, a nie wariancję opartą na genach występującą w protistycznych mtgenomach [7]. Regiony międzygenowe stanowią do 5 kb długości w grzybowych mtgenomach [7].
Chociaż protistyczne mtgenomy zawierają więcej genów, grzybowe mtgenomy zawierają znacznie większą ilość genów kodujących TRNA [6,7]. Grzybowe rozmiary mtgenome obejmują większy zakres w porównaniu do protist Mtgenomes. Najmniejszy znany grzybowy mtgenome to 19 kbp, znaleziony w Schizosaccharomyces pombe [6]. Największy znany grzybowy mtgenome to 100 kbp, znaleziony w Podospora Anserina [6]. W przeciwieństwie do Protist Mtgenomes, zawartość genu grzybowego mtDNA jest stosunkowo spójna między organizmami [6].
Źródła składników odżywczych i strategie pozyskiwania składników odżywczych
Grzybowe pozyskiwanie składników odżywczych
Grzyby używają grzybni, ich kolekcji strzępek, do pozyskiwania i transportu składników odżywczych przez błonę plazmatyczną ich komórek [2]. Proces ten jest wysoce zależny od pH środowiska, od którego nabywane są składniki odżywcze [2]. Grzyby to saprotrofy, nabywając swoje składniki odżywcze przede wszystkim z rozpuszczonej materii organicznej rozkładającej się martwych roślin i zwierząt [1]. Wszelkie wymagane trawienie składników odżywczych występuje zewnątrzkomórkowo przez uwalnianie enzymów, które rozkładają składniki odżywcze na monomery, które zostaną spożyte przez ułatwioną dyfuzję [1]
Protist Acquisition
Natomiast protistów uzyskują swoje składniki odżywcze poprzez różne strategie. Próba kategoryzacji strategii akwizycji dla prototystycznych akwizycji składników odżywczych określa sześć kategorii [3]:
- Fotoutotroficzni producenci główni - Użyj światła słonecznego do syntezy składników odżywczych z CO2 i H2O.
- Bacti- & Detritivores - Żywią się bakteriami lub detrytusem.
- Saprotrofy - Żywią się pozakomórkowym trawionym, a następnie wchłoniętym materią nie żyjącą.
- Algivores - Karmij się głównie glonami.
- Nieelektywne wszystkożerniki - Karmij się nie-sekusyjnie na algach, detrytusie i bakterie.
- Drapieżniki z tytułu praporii - Karmij się przede wszystkim pierwotniakami i organizmami z wyższych poziomów troficznych.
Wiele wyżej wymienionych strategii jest mixotroficznych. Na przykład foto-autotroficzni główni producenci obejmują organizmy morskie, które mogą wykorzystać różne poziomy heterotrofii, umożliwiając pozyskiwanie składników odżywczych, które nie wymagają wkładu energii ze światła słonecznego, gdy światło słoneczne jest niedostępne [3].
Różnice reprodukcyjne
Protist i grzyby obejmują gatunki, które rozmnażają. Protisty są wyjątkowe, ponieważ obejmują organizmy zdolne zarówno do reprodukcji aeseksualnej, jak i płciowej w ciągu tego samego życia [8]. Złożoność niektórych protistycznych cykli życia powoduje oszałamiające zmiany morfologiczne w ciągu życia organizmu, co umożliwia wyraźne metody reprodukcji [8]. Zmiany morfologiczne związane z reproduktywem nie są obserwowane w grzybach, w zakresie, w jakim są w Protistach.
Aeseksualne różnice reprodukcyjne
Aeseksualne reprodukcja w grzybach odbywa się poprzez rozkład zarodników emanujących z ciał owocowych znalezionych na grzybni lub przez fragmentację grzybni lub przez pączkowanie [9]. Aesexual Reproduction u protist występuje różne metody. Rozszczepienie binarne (pojedynczy podział jądrowy) i wielokrotne rozszczepienia (wiele podziałów jądrowych) to dwie popularne metody reprodukcyjne wśród protistów [8]. Kolejną strategią reprodukcyjną specyficzną dla protisty jest plazmotomia [8]. Plazmotomia występuje wśród protistów z wielomieżnymi i pociąga za sobą podział cytoplazmatyczny bez podziału jądrowego [8].
Różnice seksualne reprodukcyjne
Rozmnażanie płciowe jest częściej wdrażane przez grzyby [8,9]. Jest również bardziej złożony niż reprodukcja eesxual, a zatem wymaga bardziej szczegółowego opisu w celu ustalenia zrozumienia, w jaki sposób proces ten różni się między protistami a grzybami.
Grzybicze reprodukcja seksualna
Podczas reprodukcji płciowej grzybowej błona jądrowa i jądro (zwykle) pozostają nienaruszone przez cały proces [9]. Plazmogamia, karyogamia i mejoza obejmują trzy sekwencyjne etapy grzybowej reprodukcji płciowej [9]. Plazmogamia pociąga za sobą fuzję protoplazmatyczną między komórkami godowymi, co wprowadza wyraźne jądra haploidalne do tej samej komórki [9]. Fuzja tych haploidalnych jąder i tworzenie jądra diploidalnego występuje w stadium kariogamii [9]. Pod koniec kariogamy i zygoty istnieje, a mejoza przebiega przez tworzenie się włókien wrzeciona w jądrze. To przywraca stan haploidalny poprzez diploidalny separacja chromosomów [9].
Strategie grzybów interakcji haploidalnych jąder podczas rozmnażania płciowego są bardziej zróżnicowane w grzybach w porównaniu z protistami. Strategie te obejmują tworzenie i wydanie gamete z Gametangia (narządy płciowe), interakcję Gametangia między dwoma organizmami i interakcją strzępek somatycznych [9].
Protist Reprodukcja płciowa
Protist Protist Secure Reproduction Strategies są prawie całkowicie odmienne z tymi, których zatrudnione przez grzyby. Strategie te pociągają za sobą unikalne procesy, które różnią się w wyniku struktury komórkowej, szczególnie przydatków komórkowych dostępnych do kontaktu z innymi protistami [8]. Formacja i uwalnianie gameta jest seksualną metodą reprodukcyjną wśród wysoce ruchliwych protystów flagowych [8]. Koniugacja jest metodą stosowaną przez Protistów CILITED, która pociąga za sobą fuzję jąder gametycznych, a nie tworzenie i uwalnianie niezależnych gamet [8]. Autogamia, proces samozapłaniania, który jest nadal uważany za formę reprodukcji płciowej, powoduje homozygotyczność wśród potomstwa samozapłodowej komórki macierzystej [8].
Podsumowanie tabeli
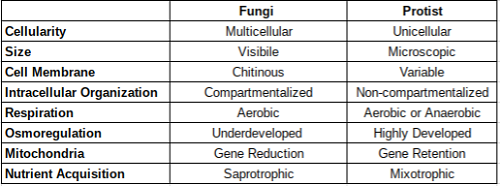
Jak podsumowano powyżej, różnice między protistami i grzybami są ogromne i można je obserwować na każdym poziomie struktury i we wszystkich ich interakcjach behawioralnych z ich środowiskiem. Ten przegląd jest jedynie podsumowaniem różnic. Cytowane referencje przedstawiają bardziej szczegółowe wyjaśnienia dla osób zainteresowanych uczeniem się więcej.